Цитоплазматическая мембрана
Содержание:
История изучения
Впервые строение и функции плазматической мембраны начали изучать в 1925 году. Тогда специалисты смогли впервые выделить оболочки эритроцитов. Они назвали их «тени», вычислили общую площадь. После этого ученые с помощью ацетона выделили все жиры (липиды). Это было необходимо для определения их количества на каждую единицу площади эритроцитов. Вывод, сделанный после исследований и экспериментов, был правильным, но ученые допустили несколько грубейших ошибок:
- ацетон не помогает выделить абсолютно все жиры из цитоплазматической мембраны;
- площадь цитолеммы была определена неправильно, поскольку мембраны были сухими.

Несмотря на эти нарушения, случайным образом результат оказался верным, что позволило открыть двойной слой или бислой. Далее исследования специалистов продолжились
Они обратили внимание на натяжение выделенных пленок. Мембраны не могли быть такими жесткими, поэтому появилась теория, что они содержат белки, позволяющие сохранять упругость и эластичность
В 1935 году американские ученые пришли к выводу, что схема строения плазматической мембраны напоминает сандвич, то есть имеется липидный бислой, с двух сторон окруженный белковыми прослойками.
В 1950-х годах теория была подтверждена во время первых микроскопических исследований. В 1960 году Дж. Робертсон сформулировал теорию строения биологической мембраны, которая утверждала, что все оболочки в клетках состоят из трех слоев. Однако теория сандвича или бутерброда была опровергнута, поскольку появились другие факты.
Первым из них стали сведения о глобулярности мембраны. Помимо этого, специалисты определили, что во время микроскопического исследования структура пленки во многом зависит от способа ее фиксации. Следующим открытием, опровергающим теорию сандвича, было изучение сперматозоида, во время которого появилось подтверждение, что даже в одной клетке структура мембраны на разных участках отличается.
Что такое клеточная мембрана
Если провести аналогию с куриным яйцом (разбив скорлупу, аккуратно отделить от нее тонкую полупрозрачную пленочку), то визуально можно представить, что скорлупа — это плотная клеточная оболочка, а пленка — мембрана. Эта картинка очень наглядно позволяет увидеть, каким образом под клеточной стенкой, состоящей из целлюлозы, располагается плазмалемма. Конечно, это представление будет условным, но, действительно, мембрана в переводе с латинского языка означает кожа. Хотя этот термин достаточно давний, он точно передает сущность мембранной структуры .
Цитолемма (еще одно ее название) животной клетки плотной оболочкой не защищена, однако имеет особый слой, состоящий из белков и жиров, соединенных с сахарами (гликопротеинов и гликолипидов). Называют его гликокаликс, и роль, которую он несет (рецепторная, сигнальная), очень важна для жизнедеятельности.
Строение
Строение структуры уникально, и именно за счет него функции клеточной мембраны выполняются точно и избирательно.
В структуру плазмалеммы входят молекулы:
- фосфолипидов,
- гликолипидов,
- холестерола,
- белков.
Однако не только такой щедрый химический состав делает цитоплазматическую мембрану особой структурой, все свои функции она выполняет благодаря строгой организации молекул.
Строение плазмалеммы физиологически идеально — двойной слой молекул жиров (липидов), полярно организованных, не дают своим выходить за пределы клетки, а чужим — проникать внутрь.
Организация плазмалеммы:
- мембрана состоит из липидов молекулы, которые имеют особое строение,
- каждый липид имеет два конца гидрофильная (любящая воду) головка и гидрофобный (боящийся воды) хвост,
- липиды выстроены таким образом, чтобы головки были снаружи, а гидрофобные хвосты внутри,
- поверхность мембраны гидрофильна (пропускает воду и, соответственно, растворы), а вот внутренняя часть, состоящая из гидрофобных окончаний, воду отталкивает,
- в основном молекулы липидов содержат остатки фосфорной кислоты (это фосфолипиды), некоторые связаны с углеводами (гликолипиды) и холестеролом,
- холестерол придает мембране упругость и жесткость,
- благодаря электростатическим свойствам липиды притягивают молекулы белков, которые также входят в структуру цитолеммы.
Именно белковые молекулы (гранулы) заслуживают отдельного внимания ученых. Из-за своего различного положения и ориентации в полужидкой липидной среде они выполняют самые различные и очень важные функции.
Внутри и на поверхности цитолеммы встречаются следующие виды белков:
- Периферические. Эти молекулы расположены на поверхности и в основном выполняют защитную и стабилизирующую функции. Так, они выстраивают ферменты в конвейерные цепи и не позволяют ферментам просто перемещаться вдоль бислоя.
- Погруженные внутрь (полуинтегральные). Основная их функция — ферментативная, также они могут участвовать в транспорте веществ. Изучена и еще одна интересная роль этих белков — как переносчиков. Они легко соединяются с транспортируемыми молекулами и проводят их внутрь клетки.
- Пронизывающие (интегральные). Они располагаются таким образом, что проходят насквозь, через билипидный слой. Если несколько таких белков сливаются, то образуется канал (пора), через которую могут проходить определенные вещества, связываясь с белковыми молекулами.
Таким образом, все элементы мембранного бислоя несут строго ограниченные своей ролью и строением функции. Благодаря такой организации система работает слаженно и точно.
Отмечено, что плазмалеммы даже внутри одной клетки неоднородны. В них различается не только соотношение химических составных (белков, липидов, углеводов), но и вязкость внутреннего содержимого, ферментативная активность, плотность наружного слоя, толщина.
Месторасположение в клетке
Мембранные структуры буквально пронизывают клеточное содержимое. Они ограничивают все органоиды (за редким исключением, например рибосомы), выстилают их изнутри, являются оболочками ядер.
Самая массивная по содержанию плазмалеммы структура — эндоплазматическая сеть (ЭПР). Если сложить все мембраны, ее составляющие, то получится площадь более половины общей — на все клеточное пространство. По морфологии оболочка ЭПР сходна с внешней ядерной. Они составляют с ней единую систему и обеспечивают активный взаимный перенос элементов.
Комплекс Гольджи — еще один органоид, полностью выполненный из мембранных мешочков (цистерн). Также цитолеммы имеют митохондрии и пластиды.
Плазматическая мембрана — это часть плазмалеммы, находящаяся на границе клеточного содержимого. Она ограничивает протопласт от внешней среды, окружает клетку, защищая его от наружного воздействия.
Функции
В зависимости от расположения и особенностей все мембраны выполняют собственные функции, тем не менее по выполняемой работе они сходны.
Роль плазмалеммы:
- Барьерная. Эта функция является основной и выполняется всеми видами клеточных мембран. Особенно она важна для наружной оболочки: благодаря ей клетка поддерживает форму, гомеостаз, стабильность внутреннего содержимого, целостность.
- Транспортная. Второе важнейшее назначение — активный и пассивный перенос веществ изнутри клетки в наружную среду и обратно. Механизмы этого переноса самые разнообразные, транспорт может происходить как через каналы, образуемые пронизывающими молекулами белков, так и с помощью переносчиков. Также различают пассивное (по градиенту концентрации, например диффузия газов), и активное (против градиента, с затратой выработанной клеткой энергии).
- Рецепторная. Эта роль возложена на пронизывающие белки, которые особым образом связаны с углеводными цепочками (гликополисахаридами). Образовавшиеся таким образом рецепторы, которые по своему строению и являются гликопротеидами, образуют комплекс с гормонами, затем активируются катализаторы, и такая система запускает механизмы поступления или вывода различных веществ.
- Обмен информацией. Способность клетки контактировать оболочками, обмениваясь друг с другом информацией сродни рецепторным реакциям. Благодаря им происходит стимуляция роста или торможения и иные физиологические процессы. Такой контакт может быть механическим (простое или замковое смыкание оболочек) и при помощи специальных образований — синапсов. Передающиеся через синапсы сигналы могут быть как механическими, так и электрическими.
- Энергетическая. Плазмалемма митохондрий и пластид (хлоропластов) отвечает за синтез аденозинтрифосфорной кислоты — аккумулятора клеточной энергии.
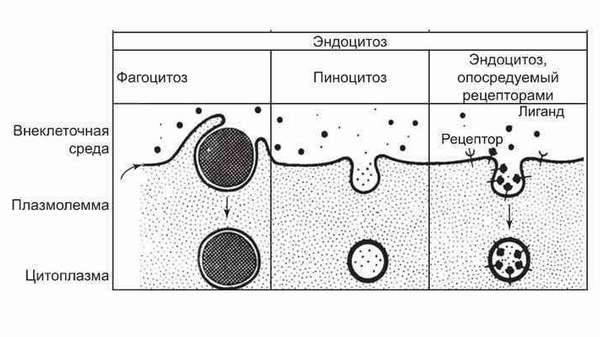
Особо следует отметить эндо- и экзоцитоз. Вследствие этих мембранных механизмов в клетку могут поступать не только целые молекулы больших размеров, но и неизмененные, сторонние клетки. Примером эндоцитоза (обволакивания крупных частиц или капель жидкости, втягивание внутрь цитоплазмы и дальнейшая химическая дезактивация) может служить поглощение вредных и чужеродных молекул лейкоцитами.
Экзоцитоз — обратный транспорт. Благодаря ему ненужные, отработанные вещества окружаются плазмалеммами и выносятся наружу через поры.
Такое множество функций и разнообразие реакций, происходящих как внутри, так и снаружи плазмалеммы, возможно за счет их упорядоченного физико-химического строения.
Клеточные оболочки
Клеточные оболочки
ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной
(синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом
(образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой
(образован целлюлозой).
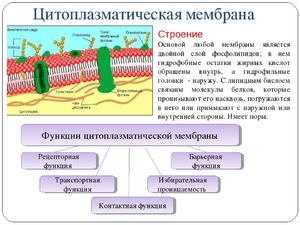
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны
. Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки
(расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки
(погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки
(пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).
А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Модели
Их существует несколько:
- «Бутербродная модель». Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- «Жидкостно-мозаичная структура». Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- «Белково-кристаллическая структура». В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.
Функции внешней стенки
Стенка бактерии призвана выполнять несколько основополагающих функций:
- быть скелетом бактерии;
- придавать ей определенную форму;
- осуществлять связь с внешней средой;
- защищать от вредных воздействий окружающих факторов;
- участвовать в делении бактериальной клетки, которая не имеет ядра и ядерной оболочки;
- удерживать на своей поверхности антигены и различного рода рецепторы (характерно для грамотрицательных бактерий).
У определенных видов бактерий есть наружная капсула, которая отличается прочностью и служит для сохранения целостности микроорганизма длительное время. В таком случае оболочка у бактерий является промежуточной формой между цитоплазмой и капсулой. Некоторые бактерии (например, лейконосток) имеют особенность заключать в одну капсулу несколько клеток. Это называется зоогелем.
Химический состав капсулы характеризуется наличием полисахаридов и большого количества воды. Капсула также может обеспечивать возможность бактерии прикрепиться к определенному объекту.
От того, насколько легко вещество проникает через оболочку, зависит степень его усвояемости бактерией. Большую вероятность проникновения имеют молекулы с длинными участками цепи, обладающими устойчивостью к биодеградации.
История исследования
В 1925 году Эверт Гортер и Франсуа Грендель (1897—1969) с помощью осмотического «удара» получили так называемые «тени» эритроцитов — их пустые оболочки. Тени сложили в стопку и определили площадь их поверхности. Затем с помощью ацетона выделили из оболочек липиды и определили количество липидов на единицу площади эритроцита — этого количества хватило на сплошной двойной слой. Хотя этот эксперимент привёл исследователей к правильному выводу, ими было допущено несколько грубых ошибок — во-первых, с помощью ацетона нельзя выделить абсолютно все липиды, а во-вторых, площадь поверхности была определена неправильно, по сухому весу. В данном случае минус на минус дал плюс, соотношение определяемых показателей случайно оказалось верным и был открыт липидный бислой.
Эксперименты с искусственными билипидными плёнками показали, что они обладают высоким поверхностным натяжением, гораздо большим, чем в клеточных мембранах. То есть в них содержится что-то, что снижает натяжение — белки. В 1935 году Джеймс Даниэлли и Хью Даусон представили научному сообществу модель «сендвича», которая говорит о том, что в основе мембраны лежит липидный бислой, по обеим сторонам от которого находятся сплошные слои белков, внутри бислоя ничего нет. Первые электронно-микроскопические исследования 1950-х годов подтвердили эту теорию — на микрофотографиях были видны 2 электронно-плотных слоя — белковые молекулы и головки липидов и один электронно-прозрачный слой между ними — хвосты липидов. Дж. Робертсон сформулировал в 1960 году теорию унитарной биологической мембраны, в которой постулировалось трёхслойное строение всех клеточных мембран.
Но постепенно накапливались аргументы против «бутербродной модели»:
- накапливались сведения о глобулярности плазматической мембраны;
- оказалось, что структура мембраны при электронной микроскопии зависит от способа её фиксации;
- плазматическая мембрана может различаться по структуре даже в одной клетке, например в головке, шейке и хвосте сперматозоида;
- «бутербродная» модель термодинамически не выгодна — для поддержания такой структуры нужно затрачивать большое количество энергии, и протащить вещество через мембрану очень сложно;
- количество белков, связанных с мембраной электростатически, очень небольшое, в основном белки очень тяжело выделить из мембраны, так как они погружены в неё.
Всё это привело к созданию в 1972 году С. Дж. Сингером и Г. Л. Николсоном жидкостно-мозаичной модели строения мембраны. Согласно этой модели белки в мембране не образуют сплошной слой на поверхности, а делятся на интегральные, полуинтегральные и периферические. Периферические белки действительно находятся на поверхности мембраны и связаны с полярными головками мембранных липидов электростатичесткими взаимодействиями, но никогда не образуют сплошной слой. Доказательствами жидкостности мембраны служат методы FRAP, FLIP и соматическая гибридизация клеток, мозаичности — метод замораживания-скалывания, при котором на сколе мембраны видны бугорки и ямки, так как белки не расщепляются, а целиком отходят в один из слоёв мембраны.
Функции
- Барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- Транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембрану обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортёры) и белки-каналы или путём эндоцитоза.При пассивном транспорте вещества пересекают липидный бислой без затрат энергии так как происходит перенос веществ из области высокой концентрации в область низкой, то есть против градиента концентрации (градиент концентрации указывает направление увеличения концентрации) путём диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.Активный транспорт требует затрат энергии, так как происходит перенос веществ из области низкой концентрации в область высокой, то есть по градиенту концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивает в клетку ионы калия (K+) и выкачивает из неё ионы натрия (Na+).
- Матричная — обеспечивает определённое взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
- Механическая — обеспечивает автономность клетки, её внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечении механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
Ультраструктура хлоропласта: 1. наружная мембрана 2. межмембранное пространство 3. внутренняя мембрана (1+2+3: оболочка) 4. строма (жидкость) 5. тилакоид с просветом (люменом) внутри 6. мембрана тилакоида 7. грана (стопка тилакоидов) 8. тилакоид (ламела) 9. зерно крахмала 10. рибосома 11. пластидная ДНК 12. пластоглобула (капля жира)
Энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки.
Рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
Ферментативная — мембранные белки нередко являются ферментами
Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
Осуществление генерации и проведения биопотенциалов.С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса.
Маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединёнными к ним разветвлёнными олигосахаридными боковыми цепями), играющие роль «антенн»
Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Геном бледной трепонемы
Так как бледная трепонема весьма важна с медицинской точки зрения, и не поддается культивированию на искусственных средах, то она стала одним из первых микроорганизмов, чей геном расшифровали исследователи. Для секвенирования был выбран штамм Nichols, выделенный в США еще в 1912 году. Геном микроорганизма представлен кольцевой двуцепочечной молекулой ДНК размером 1 138 006 пар оснований. ДНК содержит 1041 предсказанную кодирующую последовательность. Впоследствии были полностью секвенированы еще несколько штаммов T. pallidum. Ученые выяснили, что геномы штаммов отличаются, хотя и не очень значительно.
У микроорганизма идентифицировано 42 семейства генов, ответственных за основные жизнеобеспечивающие функции: механизмы репликации ДНК, транскрипции, трансляции, энергетический метаболизм, процессы клеточного деления и секреции белков.
Наличие небольшого генома с лимитированием процессов биосинтеза объясняет некоторые свойства этой бактерии. Помимо своего небольшого размера, геном бледной трепонемы имеет еще и другие своеобразные черты в виде повторяющихся генов или генов с внутренними повторами.
После расшифровки генома трепонем было выяснено, что 55% генома бледной трепонемы — это гены с предсказанной биологической ролью, 28% — ранее неизвестные гены и 17% генов не уникальны для T. pallidum, т.е. соответствуют белкам других видов бактерий.
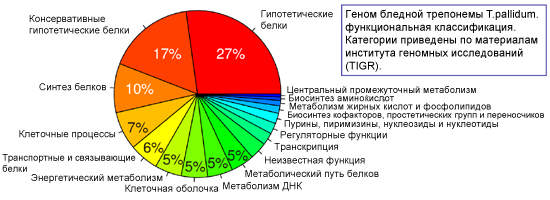
Важную роль в жизнедеятельности возбудителя сифилиса играет транспорт необходимых питательных веществ из окружающей среды. Этим объясняется присутствие широкого спектра транспортных белков с большим выбором субстратных специфичностей, кодируемых 5,7 % генома. Транспортные белки — это переносчики, связывающиеся с соответствующими субстратами внешней среды и транспортирующие их от наружной мембраны к цитоплазматической.
Как высокоспециализированный патоген, Т. pallidum не имеет в своем геноме генов, отвечающих за синтез ферментов, расщепляющих жирные кислоты, она использует сахара, содержащиеся в жидких средах организма хозяина. В качестве источников энергии микроорганизм использует глюкозу, галактозу, мальтозу и глицерин. Пути использования аминокислот как источника углерода и энергии в настоящее время не известны. Предполагают, что Т. pallidum не способна использовать аминокислоты как альтернативный источник энергии.
Одной из важнейших функций Treponema pallidum является движение, что обусловливает ее высокую инвазивность и возможность распространяться по жидкостям организма: внутрисуставной, глазной, экстрацеллюлярном матриксе и в коже. Двигательная активность обеспечивается 36 генами, кодирующими белки жгутиковых структур.