Сперматогенез — spermatogenesis
Содержание:
- Сперматогенез и оогенез
- Этапы
- Схема овогенеза и сперматогенеза
- Также в разделе
- Стадии сперматогенеза и овогенеза
- Овогенез и сперматогенез: процесс созревания
- Бластоциста
- Азооспермия. Лечение бесплодия
- Сперматогенез
- Оплодотворение
- Гормональный контроль
- Вспомогательные репродуктивные технологии (ВРТ)
- Где образуются сперматозоиды?
- Роль клеток Сертоли
Сперматогенез и оогенез
Процесс развития мужских половых клеток (сперматогенез) и женских половых клеток (оогенез) проходит ряд периодов (рис. 222). В сперматогенезе различают четыре периода: размножение, рост, созревание и формирование.
Рис. 222. Схема сперматогенеза, оогенеза и оплодотворения
Сперматогенез проходит в семенных извитых канальцах яичек, в стенке которых имеются сперматогонии (клетки полового зачатка), размножающиеся на протяжении почти всей жизни (период размножения). Часть сперматогониев, начиная с периода полового созревания, прекращает размножение и вступает в период роста, превращаясь в сперматоциты, I порядка. Эти клетки увеличиваются в размерах, и в них происходит сложная перестройка ядерного (хромосомного) вещества. В период созревания сперматоцит I порядка делится на два сперматоцита II порядка, а из последних путем нового деления образуются четыре небольших по размерам сперматиды. При этом в сперматидах оказывается одинарное число хромосом в отличие от сперматоцитов, в которых находится двойное число хромосом. В большинстве тканевых клеток человека имеются двойной набор хромосом. Мужские и женские половые клетки содержат одинарный набор хромосом. При оплодотворении происходит слияние хромосом мужской и женской половых клеток, т. е. в зиготе образуется двойной набор хромосом (см. рис. 222).
В период формирования сперматиды приобретают черты строения сперматозоидов. Появляются головка с чехликом, а также жгутик.
Оогенез проходит периоды размножения, роста и созревания. Период формирования отсутствует. Период размножения, т. е. размножение клеток полового зачатка (оогониев), заканчивается в яичниках к моменту рождения ребенка. К этому времени в яичниках имеется определенное число ооцитов I порядка (незрелых половых клеток). Начиная с периода полового созревания один раз в 4 недели, как правило, один ооцит I порядка вступает в период роста: увеличиваются в размерах его ядро и цитоплазма; в цитоплазме происходит накапливание белков, жиров и жироподобных веществ (желтка). В это же время происходит сложная перестройка ядерного вещества. Период роста сменяется периодом созревания, сущность которого сводится к делению ооцита I порядка на ооцит II порядка (крупная клетка) и очень маленькое по размерам первое редукционное тельце. Период созревания заканчивается отделением от ооцита II порядка второго редукционного тельца, тоже маленького по размерам, и появлением в результате этого зрелой женской половой клетки (яйцеклетки).
В процессе выделения первого и второго редукционных телец яйцеклетка получает одинарный набор хромосом (как и сперматиды при сперматогенезе).
Этапы
Весь процесс сперматогенеза можно разбить на несколько отдельных этапов, каждая из которых соответствует определенному типу клеток человека. В следующей таблице плоидность, число копий и количество хромосом / хроматид приведены для одной клетки, как правило, до синтеза и деления ДНК (в G1, если применимо). Первичный сперматоцит задерживается после синтеза ДНК и перед делением.
| Тип ячейки | плоидность / хромосомы у человека | Число копий ДНК / хроматиды у человека | Процесс введен ячейкой |
| сперматогоний (типы Ad, Ap и B) | диплоид (2N) / 46 | 2C / 46 | сперматоцитогенез ( митоз ) |
| первичный сперматоцит | диплоид (2N) / 46 | 4C / 2×46 | сперматидогенез (мейоз I) |
| два вторичных сперматоцита | гаплоид (N) / 23 | 2C / 2×23 | сперматидогенез (мейоз II) |
| четыре сперматиды | гаплоид (N) / 23 | С / 23 | спермиогенез |
| четыре функциональных сперматозоида | гаплоид (N) / 23 | С / 23 | семяизвержение |
Сперматоцитогенез
Процесс сперматогенеза, когда клетки переходят от первичных сперматоцитов ко вторичным сперматоцитам, сперматидам и сперматозоидам.
Цикл семенного эпителия яичка
Сперматоцитогенез является мужской формой гаметоцитогенеза и приводит к образованию сперматоцитов, обладающих половиной нормального набора генетического материала. В сперматоцитогенезе диплоидный сперматогоний , который находится в базальном отделе семенных канальцев, митотически делится, производя две диплоидные промежуточные клетки, называемые первичными сперматоцитами . Затем каждый первичный сперматоцит перемещается в адлюминальный отсек семенных канальцев и дублирует свою ДНК, а затем подвергается мейозу I с образованием двух гаплоидных вторичных сперматоцитов , которые позже снова делятся на гаплоидные сперматиды . Это деление подразумевает источники генетической изменчивости, такие как случайное включение одной из родительских хромосом и кроссовер хромосом, который увеличивает генетическую изменчивость гамет. Механизм реакции на повреждение ДНК (DDR) играет важную роль в сперматогенезе. Белок FMRP связывается с мейотическими хромосомами и регулирует динамику аппарата DDR во время сперматогенеза. FMRP, по-видимому, необходим для восстановления повреждений ДНК .
Каждое деление клетки от сперматогония до сперматиды неполное; клетки остаются связанными друг с другом мостиками цитоплазмы, что обеспечивает синхронное развитие. Не все сперматогонии делятся с образованием сперматоцитов; в противном случае запасы сперматогоний закончились бы. Вместо этого сперматогониальные стволовые клетки делятся митотически, чтобы производить копии самих себя, обеспечивая постоянный приток сперматогониев для подпитки сперматогенеза.
Сперматидогенез
Сперматидогенез — это создание сперматид из вторичных сперматоцитов. Вторичные сперматоциты, полученные ранее, быстро входят в мейоз II и делятся с образованием гаплоидных сперматид. Краткость этого этапа означает, что вторичные сперматоциты редко выявляются при гистологических исследованиях.
Спермиогенез
Во время спермиогенеза сперматиды начинают формировать хвост за счет роста микротрубочек на одной из центриолей, которая превращается в базальное тело. Эти микротрубочки образуют аксонему . Позже центриоль модифицируется в процессе . Передняя часть хвоста (называемая средней частью) утолщается, потому что митохондрии расположены вокруг аксонемы, чтобы обеспечить снабжение энергией. ДНК сперматид также подвергается упаковке, становясь сильно конденсированной. ДНК сначала упаковывается со специфическими основными белками ядра, которые впоследствии заменяются протаминами во время удлинения сперматид. Получающийся в результате плотно упакованный хроматин транскрипционно неактивен. Аппарат Гольджи окружает теперь конденсированное ядро, становясь акросомой .
Созревание происходит под действием тестостерона, который удаляет оставшуюся ненужную цитоплазму и органеллы . Избыточная цитоплазма, известная как остаточные тельца , фагоцитируется окружающими клетками Сертоли в семенниках . Полученные сперматозоиды теперь созрели, но не имеют подвижности. Зрелые сперматозоиды высвобождаются из защитных клеток Сертоли в просвет семенных канальцев в процессе, называемом спермией .
Неподвижные сперматозоиды переносятся к придатку яичка в тестикулярной жидкости, секретируемой клетками Сертоли с помощью перистальтического сокращения . Находясь в придатке яичка, сперматозоиды приобретают подвижность и становятся способными к оплодотворению. Однако транспортировка зрелых сперматозоидов через оставшуюся часть мужской репродуктивной системы достигается за счет сокращения мышц, а не недавно приобретенной подвижности сперматозоидов.
Схема овогенеза и сперматогенеза
| признаки | сперматогенез | овогенез |
| Половые железы, половые клетки | Яички, сперматозоиды | Яичники,
яйцеклетки |
| Характерные особенности половых клеток:
– относительные размеры – подвижность – форма – наличие питательных веществ |
55 мк
подвижные округлые со жгутиком отсутствует |
130-169 мк
неподвижные округлые имеется |
| Особенности развития половых клеток на разных стадиях:
– стадия размножения – стадия роста – стадия созревания |
путем митоза образуются сперматоциты;
увеличиваются в размерах; путем мейоза образуются гаплоидные сперматиды, их которых формируются сперматозоиды |
путем митоза образуются овоциты;
увеличиваются в размерах; путем мейоза образуются гаплоидные овоциты, их которых формируется яйцеклетка |
Также в разделе
| Пренатальный скрининг; хромосомные аномалии Частота врожденных пороков развития составляет 2-3%, еще 5% новорожденных имеют так называемые малые аномалии. Причинные факторы их являются гетерогенными и… | |
| Синдром частичной трисомии по короткому плечу хромосомы 9 (9р+) Синдром частичной трисомии по короткому плечу хромосомы 9 (9р+) — наиболее частая форма частичных трисомии (опубликовано около 200 сообщений о больных с такой… | |
| Синдром Хатчинсона-Гилфорда (прогерия детей) Синдром Хатчинсона-Гилфорда , или прогерия детей , — крайне редкое заболевание. Его частота составляет 1 на 1 000 000 человек. Именно этот синдром занесен под… | |
| Прогерия (преждевременное старение) Прогерия (греч. progērōs преждевременно состарившийся) — патологическое состояние, характеризующееся комплексом изменений кожи, внутренних органов,… | |
| Синдром Эдвардса (трисомия 18) Почти во всех случаях синдром Эдвардса обусловлен простой трисомнои формой (гаметическая мутация у одного из родителей). Встречаются и мозаичные формы… | |
| Клиническая диагностика наследственных болезней Большинство наследственных болезней имеет хроническое течение, вследствие чего повторная обращаемость при таких болезнях высокая. Особенно много больных с… | |
| Синдрома Вернера (прогерия взрослых) Синдрома Вернера ( прогерия взрослых) — редкое наследственное аутосомно-рецессивное соединительнотканное заболевание (М1М 272 700). Проявляется преждевременным… | |
| Синдромы частичных анеуплоидий Помимо полных трисомий и моносомий известны синдромы, связанные с частичными трисомиями и моносомиями практически по любой хромосоме. Однако эти синдромы… | |
| Синдром Марфана Синдром Марфана — наследственная доминантная болезнь соединительной ткани. Клиническая идентификация синдрома была сделана В. Марфаном в 1886 г. Причиной… | |
| Синдром Вольфа-Хиршхорна (частичная моносомия 4р-) Синдром Вольфа-Хиршхорна (частичная моносомия 4р-) — синдром, обусловленный делецией сегмента короткого плеча хромосомы 4. Клинически синдром Вольфа-Хиршхорна… |
Стадии сперматогенеза и овогенеза
На самой начальной стадии в организме человека присутствуют сперматогонии и овогонии — это предшественники сперматозоидов и ооцитов. Сперматогонии у мужчины размножаются во время всего репродуктивного возраста, их количество не ограничено. Овогонии размножаются только внутри утробы матери, а после рождения у девочки наблюдается ограниченное их количество.
На следующий стадии происходит сразу 2 этапа мейоза, то есть деление ядра клеток. Как следствие, получаются сперматиды и зрелые яйцеклетки с полярными тельцами. Из одного сперматоцита в процессе сперматогенеза появляется 4 сперматида. А из одного овоцита — 1 зрелая яйцеклетка (ооцит) и 3 полярных тельца, которые не участвуют в процессе зачатия.
Такая особенность гаметогенеза связана напрямую с функцией клеток. Сперматозоидам не нужно много питательных веществ, они должны быть маленькими и подвижными, чтобы быстро проникнуть в маточную трубу и оплодотворить женскую половую клетку. Яйцеклетке же не нужна подвижность. Она крупная и способная к делению, ей нужно много питательных веществ, чтобы после оплодотворения начать размножаться.
Как следствие, в процессе сперматогенеза формируется огромное количество сперматозоидов, а яйцеклетка в период овуляции выходит только одна, в более редких случаях две или три.
На последней стадии сперматиды продолжают развиваться, а яйцеклетка к этому моменту уже полностью созрела. Мужские гаметы принимают правильную форму и становятся сперматозоидами, это необходимо, что половые клетки могли двигаться. Длительность сперматогенеза составляет 74 дня.
Овогенез и сперматогенез: процесс созревания
Процесс подготовки гамет к оплодотворению начинается в половых железах, у мужчин данный орган представлен тестикулами, а у женщин яичниками. В целом гаметогенез у обоих полов схож при прохождении первых трех стадий. Однако после этой части период созревания приобретает свои особенности у обоих полов.
Уже на первых стадиях мужского и женского гаметогенеза можно выделить свои особенности, например, мужских гамет – сперматозоидов производится всегда намного больше, чем женских – яйцеклеток. Кроме того, мужские гаметы развиваются в течение более длительного периода, чем женские.
Бластоциста
Когда морула попадает в полость матки, через прозрачную зону с микроокружения эмбриона начинает просачиваться жидкость, которая накапливается в межклеточных пространствах внутренней клеточной массы. Постепенно межклеточные пространства сливаются и формируют единую полость — бластоцель. Клетки внутренней клеточной массы образуют эмбриобласт и локализованы на одном полюсе, тогда как клетки наружной клеточной массы или трофобласт, становятся плоскими и образуют стенку бластоцисты. Прозрачная зона исчезает, что дает возможность бластоцистам осуществить имплантацию.
Клетки трофобласта, которые локализуются над эмбриобластным полюсом бластоцисты, около шестого дня начинают врастать в эпителий эндометрия — слизистой оболочки матки. Прикрепление и инвазия трофобласта обеспечиваются действием интегринов, выделяемых клетками трофобласта, а также ламинином и фибронектином межклеточного матрикса эндометрия.
Имплантация является результатом комплексных взаимодействий трофобласта и эндометрия. К концу первой недели развития зародыш человека проходит стадию морулы, бластоцисты и начинает имплантацию в слизистую оболочку матки. Итак, имплантация происходит в конце первой недели развития.
Клинические корреляции. Аномальные зародыши обычно погибают через 2-3 нед после оплодотворения, поэтому их частоту определить трудно. Полагают, что около 50% беременностей заканчиваются самопроизвольными выкидышами, и половина из них обусловлена хромосомными аномалиями. Эти выкидыши является следствием так называемых «пренатальных фильтров», что отбраковывают аномальные эмбрионы и тем самым уменьшают количество индивидов с врожденными пороками до 2-3% вместо 12%.
При комбинации методов экстракорпорального оплодотворения и полимеразной цепной реакции (ПЦР) проводится молекулярный скрининг эмбрионов с генетическими аномалиями (преимплантационная диагностика). Для этого достаточно выделить один бластомер из эмбриона ранней стадии развития и реплификовать его ДНК для дальнейшего исследования. Программа «Геном человека» занимается изучением связи специфических генов с различными клиническими синдромами.
Азооспермия. Лечение бесплодия
Необходимо различать обструктивную азооспермию (экскреторное бесплодие) и необструктивную (секреторное бесплодие) азооспермию, либо выраженную патоспермию.
Для обструктивной азооспермии характерны нормальные размеры яичек, нормальный уровень гормонов, отсутствие клеток сперматогенеза в центрифугированном эякуляте, воспалительные заболевания половых органов, операции на органах мошонки и малого таза, а также пальпаторное определение патологии придатков и семявыносящих протоков.
При необструктивной азооспермии наблюдаются признаки гипогонадизма, единичные сперматозоиды или клетки сперматогенеза в эякуляте, гормональные сдвиги, генные и хромосомные изменения, в анамнезе — токсические влияния, орхит.
Оптимальным способом лечения обструктивной азооспермии является выполнение микрохирургического восстановления семявыносящих путей с одномоментным получением сперматозоидов из зоны, располагающейся проксимальнее обструкции.
Полученный материал может быть одновременно использован в цикле ЭКО-ICSI. При этом часть сперматозоидов подвергается криоконсервации и используется в случае безуспешной операции.
Более сложным вопросом является лечение в программе ЭКО-ICSI пациентов с необструктивными формами бесплодия. Оптимальным способом получения сперматозоидов при необструктивной азооспермии в цикле ЭКО-ICSI является билатеральная мультифокальная биопсия яичек с использованием микрохирургической техники.
Известно, что при необструктивной азооспермии может быть сохранен фокальный сперматогенез. В независимости от полученных результатов гормонального и генетического исследования даже при гипергонадотропном гипогонадизме при билатеральной мультифокальной микрохирургической биопсии возможно получение сперматозоидов (до 65%).
При необструктивной азооспермии на фоне гипогонадотропного гипогонадизма первым этапом следует провести стимулирующую терапию гонадотропинами.
При выраженной патоспермии на фоне генетических аномалий, гипогонадизма (кроме гипогонадотропного), обструктивной симптоматики, варикоцеле, отсутствии АСАТ — возможно проведение эмпирической терапии, при отсутствии эффекта (либо сразу) пациент включается в программу вспомагательных репродуктивных технологий.
При необструктивной азооспермии, азооспермии неясного генеза, патоспермии любой степени выраженности (за исключением доказанной односторонней обструкции) первым этапом необходимо исключить варикоцеле, в том числе субклиническое. Проведение стимулирующей терапии целесообразно после лечения варикоцеле.
При сочетании необструктивной (неясного генеза) патоспермии (азооспермии) и антиспермальных антител проведение стимулирующего лечения противопоказано.
При идиопатическом бесплодии возможно проведение лечения гонадотропинами или антиэстрогенами, а также витаминотерапия и растительными стимуляторами сперматогенеза со сбалансированным содержанием микроэлементов. Однако эффективность такого лечения бесплодия не превышает 5%.
- Реконструкция семявыносящих путей при обтурационной азооспермии
- Восстановление семявыносящего протока после ятрогенного повреждения
- Реконструкция семявыносящих путей при обтурационной аспермии
- Азооспермия — клинический пример
Установите Flash Player для просмотра видео.
Сперматогенез
Созревание сперматозоидов (сперматогенез), в отличие от ооцитов, начинается только в пубертатном периоде и включает процессы преобразования сперматогония в сперматозоиды. При рождении мальчика половые клетки находятся в половых шнурах яичек (маленькие светлые клетки, окруженные поддерживающими клетками Сертоли). Поддерживающие клетки Сертоли происходят из клеток поверхностного эпителия яичка, подобно фолликулярных клеток яичника.
Вскоре перед половым созреванием в половых шнурах возникает просвет, и они превращаются в семенные канальцы. В настоящее время первичные половые клетки дают начало сперматогонии, среди которых есть 2 типа: сперматогоний типа А и сперматогоний типа Б. Сперматогонии типа А подразделяются митозом и обеспечивают резерв стволовых клеток, сперматогонии типа Б дают начало первичным сперматоцитам.
При нормальных условиях некоторые клетки типа А выходят из популяции стволовых клеток, чтобы начать дальнейшие генерации сперматогония, каждая из которых является более дифференцированной по сравнению с предыдущей генерацией. После завершения последнего деления клеток типа А формируются сперматогоний типа Б, который после серии митозов превращаются в первичные сперматоциты. Первичные сперматоциты вступают в продолженную профазу (22 дня), после которого наступает быстрое завершение мейоза І с образованием вторичных сперматоцитов.
Вторичные сперматоциты сразу начинают второе мейотическое разделение, в результате чего образуются гаплоидные сперматиды. В течение этих процессов, начиная с момента, когда клетки типа А оставляют популяцию стволовых клеток, и к образованию сперматида, цитокинез не заканчивается, и клетки последовательных генераций остаются связанными цитоплазматическими мостиками. Итак, потомки одного сперматогонического типа А формируют кластер половых клеток, сохраняющих контакт между собой на протяжении всей дифференциации.
Кроме того, сперматогоний и сперматиды протяжении своего развития погружены в глубокие карманы из клеток Сертоли. Клетки Сертоли обеспечивают поддержку для созревающих половых клеток, их защиту и питание, создают условия для высвобождения зрелых сперматозоидов.
Спермиогенез — ряд изменений, в результате которых сперматиды превращаются в сперматозоиды. У человека процесс преобразования сперматогоний в зрелый сперматозоид продолжается 64 суток. Спермиогенез включает следующие фазы:
1) формирование акросомы, покрывающей половину поверхности ядра и содержащей энзимы, помогающие пенетрации в яйцеклетку через ее микроокружение во время оплодотворения;
2) конденсацию ядра;
3) формирование шейки, средней части и хвоста;
4) отторжение большей части цитоплазмы.
Сформированные сперматозоиды направляются в просвет семенных канальцев. С семенных канальцев путем сокращения элементов их стенки сперматозоиды продвигаются в канальцы придатка яичка. Сначала сперматозоиды малоподвижны и приобретают активную подвижность в придатке яичка.
Клинические корреляции. Аномальные гаметы. У человека, как и у большинства млекопитающих, один фолликул яичника иногда может содержать 2 или 3 первичных ооцита. Такие ооциты могут дать начало двойни или тройни, но обычно они дегенерируют, не достигнув стадии зрелости. В редких случаях один ооцит может содержать два или три ядра. Такие многоядерные ооциты также погибают, не достигнув зрелости.
В отличие от ооцитов, аномалии сперматозоидов встречаются часто (около 10% сперматозоидов имеют дефекты). Головка, хвост, сперматозоиды могут быть маленькими, гигантскими или соединенными. Сперматозоиды с отклонениями в морфологии не имеют нормальной подвижности и обычно не способны к оплодотворению ооцитов.
Оплодотворение
Оплодотворение — это процесс слияния ядер сперматозоида и яйцеклетки и восстановление диплоидного набора хромосом. Оплодотворенная яйцеклетка носит название зиготы. Образование зиготы происходит только при проникновении сперматозоида в яйцеклетку.
Процесс оплодотворения
Этот процесс у разных организмов осуществляется неодинаково. У млекопитающих проникновение сперматозоида в яйцеклетку сопровождается растворением ее оболочки при помощи различных ферментов, выделяемых сперматозоидом. У многих насекомых яйцеклетки имеют плотную оболочку, и сперматозоид проникает через небольшие отверстия. У некоторых водных организмов на поверхности яйцеклетки образуется в месте контакта со сперматозоидом небольшой воспринимающий бугорок, который затем втягивается внутрь вместе со сперматозоидом.
Обычно в цитоплазму яйцеклетки проникает только головка сперматозоида с митохондрией и центриолью, а хвост остается снаружи. Оболочка головки растворяется, ядро начинает набухать, пока не достигнет размеров ядра яйцеклетки. Затем оба ядра сближаются и, наконец, сливаются.
Иногда в яйцеклетку одновременно проникает несколько сперматозоидов, но слияние с ядром происходит только у одного из них. В зиготе все хромосомы становятся парными: в каждой паре гомологичных хромосом одна хромосома принадлежит яйцеклетке, вторая — сперматозоиду. Это явление имеет большое значение для эволюции. Организм, развивающийся из зиготы, обладает большим диапазоном комбинативной изменчивости, следовательно и более широкими возможностями приспособления к меняющимся условиям внешней среды.
Двойное оплодотворение характерно для цветковых растений.
Гормональный контроль
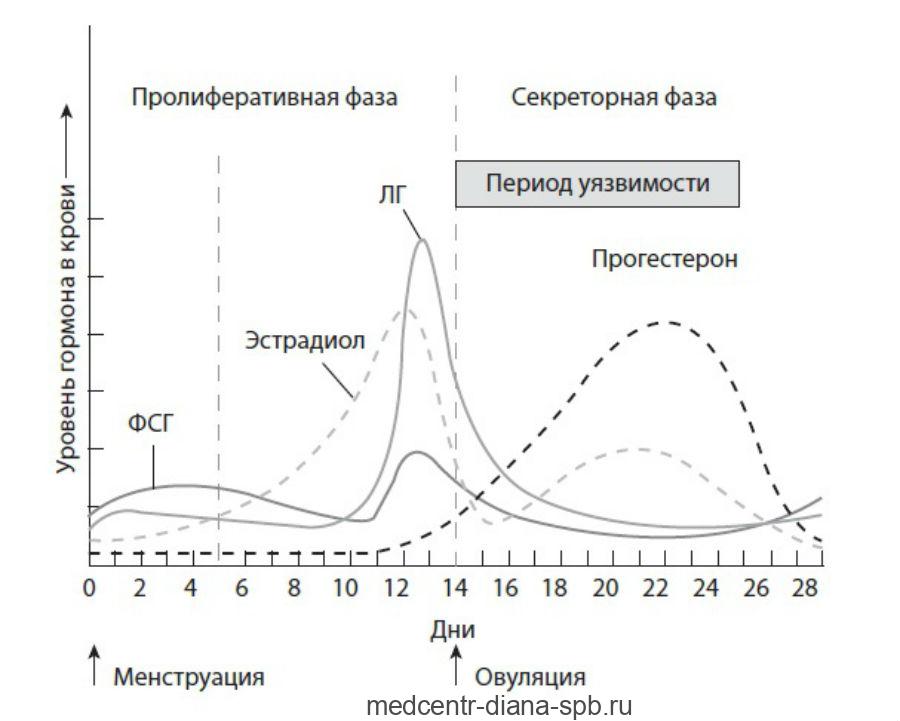
Гормональный контроль сперматогенеза зависит от вида. У людей механизм до конца не изучен; однако известно, что начало сперматогенеза происходит в период полового созревания из-за взаимодействия гипоталамуса , гипофиза и клеток Лейдига . Если гипофиз удален, сперматогенез может быть инициирован фолликулостимулирующим гормоном (ФСГ) и тестостероном . В отличие от ФСГ, лютеинизирующий гормон (ЛГ), по-видимому, играет небольшую роль в сперматогенезе, кроме индукции выработки тестостерона гонадными железами.
ФСГ стимулирует как производство андрогенсвязывающего белка (ABP) клетками Сертоли , так и образование гемато-яичкового барьера . ABP необходимо для концентрации тестостерона на достаточно высоком уровне, чтобы инициировать и поддерживать сперматогенез. Уровни интратестикулярного тестостерона в 20–100 или 50–200 раз превышают его концентрацию в крови, хотя среди здоровых мужчин наблюдаются колебания в 5–10 раз. ФСГ может инициировать секвестрацию тестостерона в яичках, но после его выработки для поддержания сперматогенеза требуется только тестостерон. Однако увеличение уровней FSH будет увеличить производство сперматозоидов, предотвращая апоптоз от типа А сперматогониев . Гормон ингибин снижает уровень ФСГ. Исследования на моделях грызунов предполагают, что гонадотропины (как ЛГ, так и ФСГ) поддерживают процесс сперматогенеза, подавляя проапоптотические сигналы и, следовательно, способствуют выживанию сперматогенных клеток.
Сами клетки Сертоли опосредуют части сперматогенеза посредством выработки гормонов. Они способны вырабатывать гормоны эстрадиол и ингибин. Клетки Лейдига также способны производить эстрадиол в дополнение к своему основному продукту — тестостерону. Было обнаружено, что эстроген необходим для сперматогенеза у животных. Однако было обнаружено, что мужчина с синдромом нечувствительности к эстрогенам (дефектный ERα ) производит сперму с нормальным количеством сперматозоидов , хотя и с аномально низкой жизнеспособностью сперматозоидов ; был ли он бесплоден, неясно. Слишком высокий уровень эстрогена может быть вредным для сперматогенеза из-за подавления секреции гонадотропина и увеличения выработки тестостерона в организме. Пролактин также важен для сперматогенеза.
Вспомогательные репродуктивные технологии (ВРТ)
Под вспомогательными репродуктивными технологиями подразумеваются:
- Внутриматочная инсеминация спермой мужа.
- Экстракорпоральное оплодотворение.
- Экстракорпоральное оплодотворение с микроманипуляцией ICSI (ИКСИ)
- Использование спермы донора во ВРТ.
Введение сперматозоидов непосредственно в полость матки предусматривает искусственное прохождение шейки матки сперматозоидами, которые могли бы погибнуть при цервикальном факторе бесплодия при попытки е стественного зачатия.
Показаниями к внутриматочной инсеминации также являются эякуляторная дисфункция, иммунологическое бесплодие, олигоастенозооспермия.
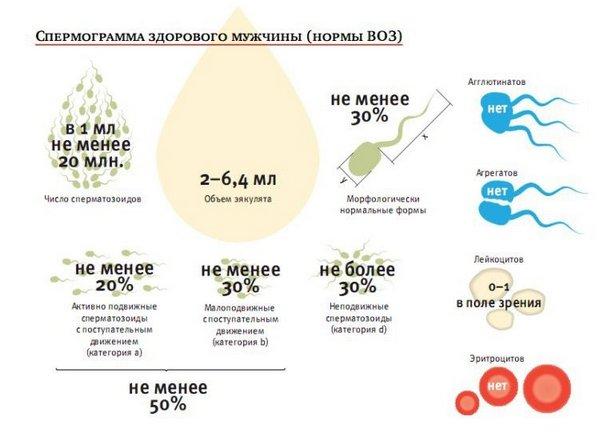
Иммунологический фактор бесплодия преодолевается путем предварительной обработки сперматозоидов. В случае неизвестной этиологии бесплодия также возможно искусственное оплодотворение методом внутриматочной инсеминации при удовлетворительных показателях качества эякулята (количество подвижных сперматозоидов более 5 млн.). Инсеминация спермы донора позволяет избежать непосредственного полового акта.
Методика экстракорпорального оплодотворения, предусматривает стимуляцию суперовуляции, что может вызвать ряд осложнений, как непосредственно при стимуляции, так вследствие многоплодной беременности. Успех процедуры стимуляции в большой степени зависит от состояния эндокринной системы женщины. Известно, что частота наступления беременности выше в тех случаях, когда при пункции фолликулов удается аспирировать более 5 яйцеклеток. Далее ооциты аспирируются путем трансвагинальной пункции яичников. После этого происходит оплодотворение in vitro — экспозиция яйцеклеток в одной среде с обработанными сперматозоидами. Затем следует перенос эмбрионов в полость матки и редукция части эмбрионов.
Возможности коррекции азооспермии оставались минимальными до внедрения методики инъекции сперматозоида в цитоплазму зрелой яйцеклетки (ICSI), требующей для оплодотворения лишь единичных сперматозоидов эпидидимального или тестикулярного происхождения.
Показаниями к ЭКО (ICSI) является: концентрация сперматозоидов ниже 10 млн. (2 млн. для ИКСИ), подвижность сперматозоидов 2 — меньше 10% (5% для ICSI), более 70% патологических форм сперматозоидов (более 96 для ИКСИ), возможность получения сперматозоидов только хирургическим путем, безуспешность других методов лечения.
Эффективность методов искусственного оплодотворения составляет около 30% за одну попытку.
Применение вспомогательных репродуктивных технологий значительно расширяет возможности коррекции наиболее тяжелых форм мужского бесплодия, включая азооспермию. Грамотное и эффективное лечение бесплодного брака возможно только при осуществлении четкого взаимодействия между урологами, гинекологами и специалистами ЭКО. Обязательным является полное информирование пациента обо всех возможных методах восстановления фертильности с указанием процента успешных мероприятий и риска развития осложнений у него самого и его супруги.
Где образуются сперматозоиды?
Для образования и созревания сперматозоида требуется 72 дня.
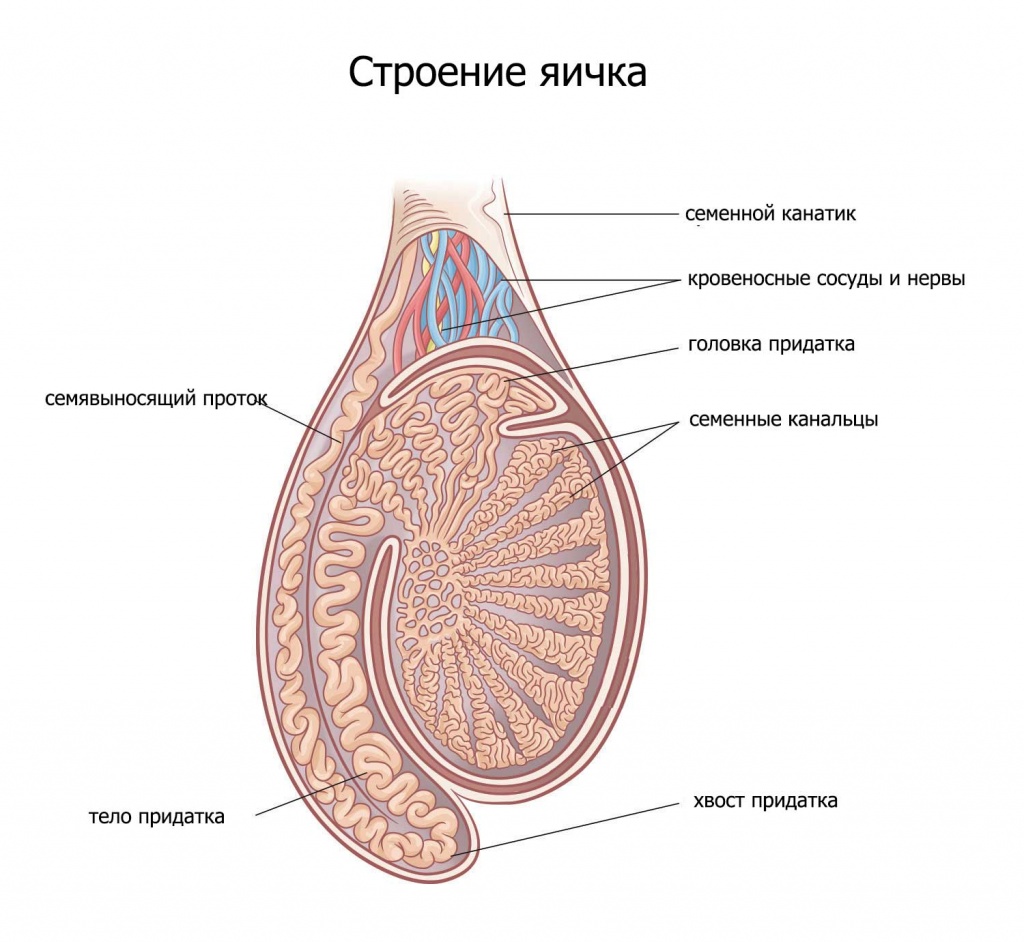
В яичках есть целая система крошечных трубок — извитых семенных канальцев. Эти трубки заполнены зародышевыми круглыми клетками, которые под действием тестостерона начинают делиться и превращаться в сперматозоиды прямо внутри стенки такой трубки. Всё это время их питают и защищают клетки Сертоли. Как только у сперматозоида формируется голова и хвост (жгутик), он проходит сквозь клетки Сертоли (гемато-тестикулярный барьер) и попадает в просвет трубки (канальца) и уже по нему начинает движение к придатку (эпидидимис).
В придатках ( а их у мужчины 2 — по одному у каждого яичка) сперматозоиды накапливаются и хранятся в течении почти 5 недель.
Параллельно с этим, предстательная железа и семенные пузырьки производят семенную плазму — беловатую жидкость, с которой смешиваются сперматозоиды и плавают в ней, попадая в семявыносящие протоки непосредственно перед эякуляцией.
Мужчина возбуждается, стимулирует половой член (мастурбация или половой акт), мышцы члена сжимаются и проталкивают эту смесь в уретру. Так мужчина эякулирует спермой — т.е. смесью семенной плазмы и сперматозоидов. В каждой порции спермы может быть до 500 миллионов сперматозоидов.
Роль клеток Сертоли
Обозначенная диаграмма организации клеток Сертоли (красный цвет) и сперматоцитов (синий цвет) в семеннике. Сперматиды, которые еще не прошли спермиевание, прикрепляются к верхушке просвета клетки.
На всех стадиях дифференцировки сперматогенные клетки находятся в тесном контакте с клетками Сертоли, которые, как считается, обеспечивают структурную и метаболическую поддержку развивающихся сперматозоидов. Одиночная клетка Сертоли простирается от базальной мембраны до просвета семенного канальца, хотя цитоплазматические отростки трудно различить на уровне световой микроскопии.
Клетки Сертоли выполняют ряд функций во время сперматогенеза, они поддерживают развивающиеся гаметы следующим образом:
- Поддерживать среду, необходимую для развития и созревания, через гемато-яичковый барьер
- Секретные вещества, инициирующие мейоз
- Секрет поддерживающей тестикулярной жидкости
- Выделяет андроген-связывающий белок (ABP), который концентрирует тестостерон
Тестостерон необходим в очень больших количествах для поддержания репродуктивного тракта, а АД обеспечивает гораздо более высокий уровень фертильности.
в непосредственной близости от развивающихся гамет.
- Секретные гормоны, влияющие на контроль гипофиза над сперматогенезом, особенно полипептидный гормон ингибин
- Остаточная цитоплазма фагоцитоза, оставшаяся после спермиогенеза
- Секреция антимюллерова гормона вызывает ухудшение мюллерова протока.
- Защищают сперматиды от иммунной системы мужчины через барьер кровь-яички
- Вклад в нишу сперматогониальных стволовых клеток
В межклеточной адгезии ICAM-1 и растворимым ICAM-1 , имеют антагонистические эффекты на плотных соединений , образующих гематоэнцефалический барьер семенников. Молекулы ICAM-2 регулируют адгезию сперматид на апикальной стороне барьера (по направлению к просвету ).